
L’eterosi, o vigore ibrido, è il fenomeno per cui un ibrido F₁ mostra caratteristiche fenotipiche superiori rispetto a entrambe le linee parentali. Questo vantaggio può manifestarsi in termini di crescita, produttività, resistenza agli stress e altri tratti biologici. I meccanismi genetici alla base dell’eterosi sono molteplici e spesso agiscono in combinazione tra loro.
Incrociando due discendenze diverse e lontane fra loro a volte si assiste a un fenomeno opposto a quello che si verifica in altri casi. Un esempio può chiarire la situazione
- Linea A 1,80 metri di altezza
- Linea B 1,50 metri di altezza
Ci si aspetta che il risultato dell’incrocio siano individui con l’altezza media di 1,65 m e questo in genere è ciò che accade.

Quando sono stati costituiti i primi ibridi commerciali di mais linee di 1,20,-1,30 metri di altezza incrociate fra loro davano origine a super piante di 4 metri di altezza.
Come è stato ottenuto queto risultato per certi versi strabiliante e chiamato “eterosi dell’ibrido”?
Si tratta di una evidente differenza di capacità produttiva, generata da individui molto scarsi per diversi aspetti del loro fenotipo. Questo aumento di “bellezza” anche estetica degli ibridi è stato chiamato in passato “lussureggiamento dell’ibrido”.
L’eterosi tra indivudui distanti

In particolare, l’eterosi o vigore ibrido si può considerare come il contrapposto a ciò che accade quando si incrociano individui strettamente imparentati fra loro, fatto che si associa al fenomeno del deterioramento del vigore dei discendenti, chiamato depressione da inbreeding, ossia depressione da auto incrocio o inincrocio. Negli esseri umani il fenomeno è riconosciuto e limitato anche dalla legge, per esempio impedendo i matrimoni fra fratelli, sorelle e anche cugini di primo grado. Questa situazione, comunque, si manifesta anche tra cugini di secondo grado e in linea generale in comunità di popolazioni isolate e ridotte di numero. Questo fatto costringe gli individui a doversi riprodurre con consanguinei. Un esempio tipico è la situazione che si crea tra le popolazioni di isole o di valli difficili da raggiungere, dove avvengono incroci unicamente tra individui appartenenti alla stessa comunità locale.
Si ipotizzi un incrocio tra due individui, appartenenti sia a specie animali che vegetali. Per esempio un individuo maschio e un individuo femmina di altezza pari a 1,60 e 1,70 cm. Trascurando le differenze dovute al dimorfismo sessuale, ad esempio nella specie umana le donne sono significativamente più basse degli uomini, ci aspettiamo che nasca un individuo di 1,65 metri di altezza, ossia la media dell’altezza dei genotipi parentali. Questo è ciò che accade nella maggior parte dei casi. È normale che la discendenza di due individui con caratteri quantitativi diversi si collochi intorno alla media dei due parentali. I caratteri quantitativi sono i caratteri “misurabili”, come l’altezza, la massa, la “riproduttività” ossia la capacità di lasciare un numero più o meno elevato di discendenti che spesso costituisce anche un parametro economico e in tal caso si parla di “produttività”, la lunghezza delle foglie, la superficie fogliare e così via. I caratteri qualitativi invece si manifestano come caratteri alternativi opposti e non sono misurati, esempio: rosso e bianco, verde o giallo, inserzione del seme, rampicante e non rampicante.
Caratteri Quantitativi: Media e Valore Atteso della Discendenza
Un carattere misurabile può essere espresso come misura del suo valore in funzione della sua variabilità nella popolazione. Esistono parametri statistici che sono in grado di misurare sia la media che la dispersione intorno alla media.
Pertanto, si prevede ragionevolmente che il valore atteso di una discendenza sia il valore medio del carattere negli individui parentali.
Possibili spiegazioni dell’eterosi
Gli studi successivi alla riscoperta delle leggi di Mendel
Subito dopo la riscoperta delle leggi di Mendel, a inizio del XX secolo, negli Stati Uniti partirono una serie di ricerche scientifiche di tipo applicativo, volte a migliorare la produttività del mais. La produttività del mais consiste nella capacità del mais di produrre da 600 a 1000 semi e più da un singolo seme. Sostanzialmente una sola pianta di mais produce 1000 figli e può teoricamente dare luogo a una discendenza di 1000 individui. Ciò rende il mais la pianta più ri-produttiva del mondo e una specie relativamente facile da indagare e da studiare con metodi statistici, dando origine a discendenze numerosissime in cui i parametri della media e della deviazione standard sono calcolati a partire da campioni di popolazione molto estesi.
Fu subito evidente che i metodi di selezione usati a quel tempo erano totalmente sbagliati e non davano alcuna risposta desiderata. In pratica il seme scelto per la riproduzione rispondeva a criteri estetici o a criteri ritenuti validi dal punto di vista agronomico che si rivelarono totalmente errati, una volta sottoposti a incroci di tipo mendeliano mirati a misurare l’ereditarietà dei caratteri favorevoli, i quali hanno come effetto elevate produzioni.
Risultati con il mais
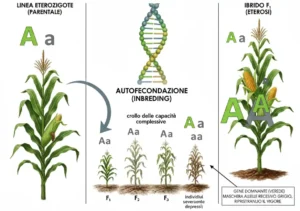
Il mais, a differenza dei piselli studiati da Mendel, è allogamo e non autogamo, e per studiare in modo corretto la discendenza è necessario effettuare una serie di incroci mirati a sostituire la fecondazione incrociata con l’autofecondazione. Proprio per questo, esattamente come accade per gli esseri umani e per gli animali, anche le piante allogame sottoposte ad autofecondazioni ripetute sono soggette alla “depressione” da inbreeding, ossia alla comparsa di difetti genetici e a un generale deperimento delle piante.
La discendenza di un individuo è una linea genetica, ad esempio tutti i nostri figli e i nostri nipoti e i nostri bisnipoti sono una linea genetica. Nel mais una sola pianta può dare origine a centinaia di discendenti e ognuno di essi può essere autofecondato. Si procede in questo modo a formare una linea genetica derivante da un individuo a cui possiamo dare un nome , in base al luogo di origine, al nome dello scienziato che ha effettuato gli incroci, al nome della località in cui viene effettuato l’esperimento o a un nome di fantasia dato dal costitutore, talvolta in onore di qualche personaggio famoso o della donna amata.
Meccanismi principali dell’eterosi
-
Ipotesi della Dominanza
Secondo questa ipotesi, i geni dominanti degli ibridi mascherano gli alleli recessivi svantaggiosi provenienti da ciascun genitore, migliorando il fenotipo complessivo.
Supponiamo di autoincrociare delle linee di piante eterozigoti, ossia di piante che naturalmente si incrociano fra loro tra piante diverse. Se incrociamo la pianta con sé stessa si assiste a un crollo delle capacità complessive del genotipo e tutti i parametri diminuiscono di intensità. Ad esempio l’altezza diminuirà considerevolmente già a partire dalla prima generazione di autoincrocio e continuerà a diminuire negli incroci successivi.
Cosa è accaduto? Per l’ipotesi della dominanza l’autoincrocio aumenta il numero di omozigoti, sia dominanti che recessivi, mentre diminuisce il numero di eterozigoti. Nella popolazione, inevitabilmente, si fissano genotipi omozigoti e una quota dei geni diventerà omozigote.
Genotipo parentale di partenza: AaBbCcDd
Dall’autoincrocio in prima generazione derivano 256 possibili linee. Tra esse aumenterà la frequenza di genotipi omozigoti. Se i parentali sono eterozigoti per tutti i geni, i discendenti di prima generazione porteranno la percentuale di genotipi omozigoti per ogni gene al 50%
Si formeranno sia genotipi AABBCCDD, che genotipi aabbccdd.
In seconda generazione la percentuale di omozigoti sale al 75%, in terza generazione la percentuale di omozigoti sale all’87,5%, in quarta generazione al 93,75%, per andare al 96,875% in quinta generazione.
Il concetto di linea pura
Una linea genetica è la discendenza di un solo individuo, alla quinta generazione di autoincroci si considera la linea genetica una linea pura, ossia sostanzialmente con tutti i geni presenti in forma omozigote (AA o aa).
L’autoincrocio ha la caratteristica di mettere in risalto tutti i difetti coperti dalla presenza di un allele funzionante e i successivi autoincroci portano alla formazione di individui severamente depressi per la presenza di numerosi alleli omozigoti recessivi che non svolgono le funzioni vitali di cui sono responsabili.
Perché fare questo se si ottengono individui così severamente penalizzati? Alcuni individui delle linee pure, nonostante la loro bassa produttività e altezza sono in ogni caso migliori di altri. Le linee con “meno difetti”, ossia i discendenti delle piante migliori, che conservano comunque una capacità vitale, possono essere scelte come linee da cui partire per i successivi autoincroci.
A un certo punto cambia l’impostazione dell’esperimento. Le miglior linee pure, la cui riproduzione è stata separata fino alla quinta generazione, sono di nuovo incrociate fra loro, ripristinando in questo modo gli eterozigoti.
Combinazioni che portano all’eterosi secondo l’ipotesi della dominanza

Non tutti gli ibridi che nascono in questo modo manifestano caratteri superiori ai parentali, ossia alle varietà originarie da cui sono state tratte le linee pure.
Questo perché per una combinazione ottimale è necessario che ABCde, si incontri con abcDE, e quindi tutti i geni siano di nuovo funzionanti in un ibrido che presenta tutti gli alleli ABCDE.
Se le linee che si incrociano hanno gli stessi geni omozigoti o mancano di intere sequenze di alleli che sono necessari allo sviluppo normale della pianta l’ibrido non funziona come previsto dalla teoria della dominanza.
Storicamente sono stati necessari trent’anni per formare i primi ibridi commerciali, ma da allora la maggior parte delle sementi commerciali è ibrida. Nel tempo, inoltre, le linee dopo decenni di selezione sono state sempre più arricchite di geni favorevoli e gli incroci fra loro si sono dimostrati sempre più validi dal punto di vista produttivo e in generale per tutte le caratteristiche favorevoli alla coltivazione, ad esempio la resistenza a numerose malattie.
-
Ipotesi della Superdominanza
La superdominanza (o eterozigosi vantaggiosa) postula che in alcuni loci l’eterozigote possiede un vantaggio intrinseco rispetto a entrambi gli omozigoti. Ciò non dipende dalla semplice mascheratura di alleli negativi, ma da una reale superiorità funzionale dell’eterozigote.
Un esempio biologico utile per spiegare la superdominanza riguarda geni che codificano per enzimi con diverse temperature ottimali di lavoro. Supponiamo che:
– l’allele A produca un enzima con un ottimo di attività a 25 °C,
– l’allele A1 produca un enzima simile, ma mutato e con attività ottimale a 15 °C.
Un individuo eterozigote AA1 può esprimere entrambi gli enzimi, risultando efficiente su un intervallo di temperatura più ampio (es. 15–25 °C), mentre gli omozigoti AA o A1A1 avrebbero un’efficienza limitata a una singola condizione ambientale. In questo caso, l’eterozigote ha un vantaggio concreto e autonomo, non dipendente da alleli recessivi difettosi.
Questo esempio rappresenta una forma di complementarità funzionale, oggi studiata anche in relazione a enzimi regolatori, geni omeotici, regolazione dell’espressione genica e adattamenti ambientali complessi.
Esempi documentati di superdominanza
Anemia falciforme e resistenza alla malaria
L’esempio classico di overdominance riguarda il gene della beta-globina. Gli individui eterozigoti AS sono resistenti alla malaria e non sviluppano l’anemia falciforme, mentre gli omozigoti AA sono suscettibili alla malaria e gli SS soffrono della malattia.
- Complesso maggiore di istocompatibilità (MHC) nei pesci artici
In alcune popolazioni di salmonidi, gli eterozigoti per i geni MHC mostrano maggiore resistenza a patogeni e una fitness superiore rispetto agli omozigoti, rappresentando un chiaro caso di vantaggio eterozigote.
3. Gymnadenia rhellicani – colore dei fiori e impollinazione
Nell’orchidea alpina Gymnadenia rhellicani, gli eterozigoti tra fiori bianchi e neri producono fiori rossi intermedi, che attraggono sia api che mosche. Questo porta a una maggiore fitness riproduttiva.
- Mutazione ebony in Drosophila melanogaster
I moscerini eterozigoti per la mutazione *ebony* (e⁺/e) mostrano una sopravvivenza superiore rispetto a entrambi gli omozigoti, configurando un esempio sperimentale di superdominanza nel contesto dell’adattamento.
Tabella riassuntiva dei casi di superdominanza
- Anemia falciforme: AS > AA e SS – vantaggio immunitario
- MHC nei pesci: eterozigoti > omozigoti – ampia copertura immunitaria
- Gymnadenia: colore intermedio > estremi – attrattività per impollinatori
- Drosophila: e⁺/e > e⁺/e⁺ o e/e – migliore sopravvivenza
Superdominanza osservata in natura
Il modello ipotetico degli enzimi con differenti temperature ottimali è coerente con molti casi reali di superdominanza osservata in natura. Tali fenomeni dimostrano che gli eterozigoti possono ottenere vantaggi funzionali concreti, non solo per mascheramento di alleli difettosi, ma anche per complementarità o sinergia tra funzioni genetiche distinte. Questo concetto è centrale nella genetica evoluzionistica, nella fisiologia ambientale e nella selezione di varietà ibride.
3. Ipotesi delle Interazioni Non Alleliche (INA)
L’eterosi può anche derivare da epistasi positiva: interazioni tra geni localizzati in loci differenti. Queste interazioni, presenti solo nell’ibrido, attivano percorsi metabolici, regolazioni o reti funzionali non attive nei genitori. In particolare, alcune combinazioni di geni complementari possono portare a risultati fenotipici superiori rispetto alla somma dei singoli effetti genici (effetto sinergico).
Modello combinato
Oggi si ritiene che nessuna ipotesi escluda le altre. Piuttosto, l’ibrido F₁ beneficia della combinazione di più meccanismi: dominanza per eliminare difetti recessivi, superdominanza per esprimere vantaggi funzionali degli eterozigoti e interazioni non alleliche per sfruttare reti geniche sinergiche. L’insieme di questi fattori produce un fenotipo più robusto, adattabile e produttivo.
4 Nuove ipotesi: Effetti Pleiotropici e Loro Ruolo nell’Eterosi
Cos’è la pleiotropia?
La pleiotropia è il fenomeno per cui un singolo gene influenza più di un carattere fenotipico. In altre parole, un gene non ha un solo effetto, ma diversi effetti su tratti distinti dell’organismo.
Esempio classico:
- Il gene della fenilchetonuria (PKU), se mutato, causa:
accumulo di fenilalanina (difetto metabolico), - ritardo mentale,
- alterazioni della pigmentazione della pelle,
- problemi motori e neurologici.
Tutti questi effetti derivano da un’unica mutazione: questo è pleiotropismo.
Tipi di pleiotropia
- Pleiotropia molecolare diretta
Il prodotto del gene (es. una proteina) agisce in diversi tessuti o organi con funzioni multiple.
- Pleiotropia secondaria o causale
Un effetto primario del gene innesca cascate metaboliche che influenzano più caratteri.
- Pleiotropia dello sviluppo
Un gene coinvolto nelle fasi precoci dello sviluppo (es. geni omeotici) condiziona l’intera morfologia o fisiologia dell’organismo.
Pleiotropia ed eterosi
Nell’ambito dell’eterosi, gli effetti pleiotropici possono contribuire in modo significativo al vantaggio dell’ibrido. Quando un gene eterozigote esercita effetti positivi multipli, il suo contributo complessivo al fenotipo sarà maggiore di quello previsto da singoli caratteri. Questo vale sia nei casi di dominanza (mascheramento di effetti negativi), sia nei casi di superdominanza (vantaggi funzionali propri dell’eterozigote).
Esempio in agricoltura: il gene tb1 del mais
Il gene teosinte branched1 (tb1) è un esempio noto di pleiotropia:
– Regola la ramificazione della pianta (meno rami laterali),
– Aumenta la dimensione della spiga,
– Riduce l’infiorescenza maschile laterale.
Questi effetti combinati hanno permesso la domesticazione del mais da teosinte, e oggi il gene tb1 ha effetti pleiotropici favorevoli in molti ibridi, migliorando produttività e forma della pianta.
Connessione con la superdominanza
Immagina un gene con due alleli:
– A: ottimizza la crescita vegetativa
– A1: ottimizza la formazione del seme
L’eterozigote A/A1 potrebbe trarre beneficio da entrambi gli effetti, risultando superiore rispetto ad A/A o A1/A1. Qui la pleiotropia si somma alla complementarietà allelica, dando origine a una superdominanza pleiotropica.
Esempi pratici: l’epopea del mais ibrido
L’eterosi rappresenta un fenomeno complesso, centrale in agronomia e genetica applicata. La comprensione dei suoi meccanismi richiede l’integrazione di genetica classica, molecolare e genomica funzionale. Lo studio dei diversi modelli (dominanza, superdominanza, epistasi) resta fondamentale per sviluppare nuove varietà ibride più efficienti e resilienti.
Il mais (Zea mays) è uno dei casi più emblematici in cui l’eterosi ha rivoluzionato l’agricoltura moderna. Negli Stati Uniti, a partire dagli anni ’20 del Novecento, vennero costituite le prime varietà ibride combinando due linee pure (inbred lines), ciascuna ottenuta per autofecondazione prolungata (inbreeding). Tali linee erano geneticamente stabili, ma poco vigorose. Tuttavia, il loro incrocio produceva ibridi F₁ con una produttività molto superiore.
Due delle linee più celebri sono:
– **Stiff Stalk Synthetic (SS)**: sviluppata presso l’Università dell’Iowa, è una linea altamente omozigote, caratterizzata da fusti rigidi, resistenza all’allettamento e buona fertilità maschile.
– **Lancaster Sure Crop (LSC)**: originaria della Pennsylvania, presenta buona resistenza a malattie e adattabilità ad ambienti umidi.
Conclusione
L’incrocio tra linee derivate da SS e da LSC ha dato origine ad alcuni dei più importanti ibridi commerciali di mais del XX secolo. Questi ibridi mostrano un marcato vigore ibrido, con aumenti di resa fino al 30–40% rispetto ai genitori. L’eterosi in questo caso è dovuta a un insieme di fattori:
- mascheramento di alleli recessivi svantaggiosi,
- superdominanza in alcuni loci chiave (es. quelli legati all’efficienza fotosintetica e all’apparato radicale),
- interazioni epistatiche positive tra geni provenienti da genotipi distinti.
Il successo degli ibridi SS × LSC ha portato allo sviluppo di una vera e propria industria sementiera basata sugli ibridi F₁. Ancora oggi, molte aziende selezionano linee parentali in base alla loro combinabilità con Stiff Stalk e Lancaster, a conferma del valore storico e genetico di questa combinazione.






{kind=link}